- Cilia. The sucking cilia are known to be unbalanced. Although spikes can nerf them, organisms often don’t develop spikes, and even if they do, you can jump to another patch that hasn’t evolved to deal with you. They’d also be really unfun to play against, except that auto-evo doesn’t evolve them, presumably due to part additions outpacing auto-evo being updated to account for them. They also look weird in the editor, as all of their waves are synchronised. A random time delay would fix this. And finally, when you die, their effects still work, which gets really weird, because when you eject a dead microbe, or die yourself, you’re surrounded with floating parts and they fly absolutely everywhere, at ridiculous speeds.
- Variants. Variant parts are pretty interesting, but they’re not obvious (I assume they’ve been added to the tutorial, at least). You don’t have an indication on the parts screen about which parts have variants. The only three parts I know to have variants are the slime jet, pilus, and cilia, and 2 of those have variants with completely different functions from the main part. (The slime jet goes from a speed part to a defensive part, and the cilia goes from a speed part to an offensive part.) Also, I’d have to suspect that auto-evo doesn’t take them into account.
- Endosymbiosis: Almost all of the organisms I see require 5 engulfs, which is basically impossible since in 5 (or more) generations they’ll almost certainly go extinct and disappear from the patch.
- Gases. With Experimental features turned on, gases disintegrate over time and anaerobic respiration is dominant. With the feature off, the opposite happens; surface patches gain an ever-increasing amount of CO2 and oxygen. From memory, I recall multiple patches with around 45% O2 and 15% CO2, both slowly trending upwards. This is very unbalanced and makes aerobic respiration chaotic (I tried to go into deep water for an endosymbiont, but I couldn’t do it because of the drastic difference in O2 from the surface to the lower level - 2/3 less down there or something) to use.
- Digestion is bugged. This is known, I believe. Digested cells sometimes (not always) stick in you and produce chunks if you eject them. Also, it produces a lot of glucose. I consistently had a massive surplus of glucose, although that might be because of #1 and how easy it made hunting. I could easily gather several eukaryotes in a very short hunt, there was no real challenge. Also, for some reason, my cell was able to acquire a giant iron chunk (no rusticyanin so it didn’t stick), but was too small to eat a cell of similar size (around 50-60 IIRC). I’d have to guess that this is because of the dozen vacuoles I installed, but it would still imply that something’s mismatched.
1 Like
Would you count flagellum length changing as a variant or something else?
1 Like
So you haven’t played with the tutorials?
The first time you place an organelle that can be modified there’s a tutorial popup about modifying organelles.
Also both toxin shooter organelles can be modified to change the toxin type, chemoreceptor can be modified to set what it detects, vacuoles can be specialized for a certain compound.
Our eventual goal is that each organelle would have at least one upgrade / modification possible. That way there’s really no point in showing them in the GUI as the player should expect that they can modify anything they want for other functionality.
There’s been enough feedback on that already. The experimental compound mode is probably going to be entirely scrapped from 0.8.0 as it doesn’t look like it’s going to be worked on any further. I’m currently in the process of adding enough compound dynamism to the default mode in a balanced manner to consider dynamic compounds done.
Each eukaryotic hex counts as 2 hexes in engulf size, and many eukaryotic parts are often multiple hexes. So it is not far fetched to be big enough (especially if close to reproducing) in engulf size to eat a big iron.
1 Like
Aight.
I finished my run, did another, and I have a lot more to talk about.
- Playing as a eukaryotic apex predator is by far the easiest niche, at least in my experience (although I haven’t really done chemotroph or ferrotroph runs). It is not difficult to become the uncontested ruler of your patch, and in fact, it’s not hard to completely genocide a patch so that you kill off all of your food sources.
- This is exacerbated by the fact that atmospheric oxygenation is completely broken, with the atmosphere chaotically fluctuating. Generally, oxygen will go up while CO2 will remain stable, but CO2 can also go up. There are no lower or upper limits on either gas, nor any limits on the total gas quantities, so you can easily see a patch missing 30% of its atmosphere (denoted by “Other”), and conversely a patch whose atmospheric gases sum to 130%. Oxygenation is highly variable, meaning that a cell which can survive in one patch easily may be completely incompatible with a patch one sea-level down. But those survivable patches are very survivable - while hydrogenase doesn’t scale, metabolosomes and especially mitochondria scale to the extreme, with singular mitochondria producing massive amounts of ATP. I’m not sure of the exact number, but the patches I inhabited in the end stages of my second run had around 40% oxygen, I believe, and I’d hazard a guess that each mitochondrion was producing 15-20 ATP.
To further explain this - I found an estuary patch with 0.5% oxygen and 43% CO2, which put it above 100% atmosphere since it had 60% Nitrogen. Its neighboring patches in its seaside cluster were both 2.05% oxygen and 2.5% CO2, with a 35.4% hole in the atmosphere as a result. None of this arrangement is working correctly. After one generation, this estuary of 43% CO2 crashed to 3.23% oxygen and 8.85% CO2, thus matching its neighbors with a 28% atmospheric deficit. The two neighboring patches did not improve their deficits much, but gained in both CO2 and oxygen. I suspect the root cause of this glitch is some error with interpatch gas exchange, assuming that’s a thing. - Sea patch clusters can be variable, but usually they look pretty similar to one another. Seaside patch clusters almost always look the same. One of each type, in a triangular arrangement.
- When you’re bound to other cells, or you’ve become multicellular, I observe a phenomenon wherein your cell becomes very fast, but no matter how many cilia you collectively have, you effectively cannot turn, heavily limiting your mobility and hunting efficacy. As well, when you engulf while bound to other cells, the compound sharing glitches out, and if you unbind, you can retain Ingested Matter even if you do not actually have any in your cell. If I remember correctly, you retain the full quantity of IM, not even divvied up amongst the cells, but I’m unsure of that. Also, there’s no reason for your cell to bind to other cells if you aren’t planning to jump to multicellularity, which breaks the graduality of evolution. (I’d suggest binding being useful for genetic exchange (for reduced costs and/or free parts in the editor?) as a way to justify evolving it when you aren’t planning to multicellular immediately.)
- In my first run, I don’t believe I received a ‘You Have Thrived’ message even though I very well did. Not very relevant.
- Auto-evo has replaced the axehead cells with several new varieties of artificially intelligent seizure. Firstly, I observed a staggering quantity of cells which decided to spontaneously evolve vertical chains of 3-5 hydrogenases, and on occasion, they would do this several times over multiple steps. This didn’t seem to be particularly disadvantageous. I also saw this occur with metabolosomes (that evolution killed my endosymbiont at the time). Secondly, auto-evo seems to now adore the designs of a single hex of cytoplasm, a single hex of cytoplasm with one food protein directly below it, and a single hex of cytoplasm with two distinct food proteins directly below it. These evolve repeatedly amongst many unrelated species. Thirdly, auto-evo repeatedly evolves the same cells. For an extreme example, I saw two cells which diverged from a common ancestor, and were identical. They both matched the layout of one cyto, one food (specifically a rusticyanin). They evolved the next generation, changing literally nothing. The next generation, they both evolved to become mono-cytoplasm. One of the two diverged another, a species which was… a single hex of cytoplasm. So now there are three which look identical. Auto-evo might be considering color changes as valid reasons to diverge species. Similarly, there was a species which consisted of one cytoplasm, one rusticyanin, and one sulfur protein (forgot what those were called). This evolved to lose its rusticyanin. A species offshoot retained the rusticyanin. This should be reversed. The new species should be the one losing the rusticyanin in this arrangement. This problem remains prevalent throughout the game, but is of course most obvious in earlygame, as cells haven’t become very complex.
- Endosymbiosis is basically irrelevant. Most endosymbionts are apparently unsuitable and require 4 or 5 engulfs to acquire; auto-evo additionally seems to adore cells that do not metabolise glucose, since I almost never saw mitochondrion candidates. There were some chloroplasts, but most of the time, I’d see ferroplasts or chemoplasts - even though I do not play in volcanic vents! It’s a cool idea, but it doesn’t seem to work very well right now.
- For UI issues, the variation screen seems to imply room for 8 variants of given parts, which is definitely not planned, so that should be shrank. When you examine the [?] for eukaryotic organelles, there’s so many not unlocked at the start of the game that they go off the screen. The tutorial suggests that the player remain a single hex of cytoplasm until reaching the surface and evolving thylakoids, but a new player who tries this might get confused when the editor tutorials tell them that they should always be using as much MP as possible in the editor, since it’s not conserved. The change of the color of engulfing between bright and dark blue is never explained, so I don’t know what it signifies. The chemoreceptor UI could also do with some improvements, as the meanings of the two sliders are about as clear as concrete. I especially take issue with the ‘patch size’ slider. I have no idea how it works or how it judges what the size of a patch is, so it should definitely be improved in that respect.
- The high MP cost of Hard difficulty is really unfun to me. Because the prices of most parts are just over 50, you can effectively only make one addition to your cell most edits. This seems like it would make progression really difficult and slow. It might be less extreme of a limitation with more variants being added, as that might permit more alterations to your cell that are less expensive than a new part.
- Although marine snow is perfectly sensible later on, in the earlygame, it’s nonsensical. You can see marine snow made of parts that have never been evolved in the world, including eukaryotic parts in a world with no eukaryotes. This is especially egregious when a player quickly moving between patches finds marine snow in a patch without any other microbes in it or the patches above it.
3 Likes
- I don’t see many eukaryotes that aren’t related to the player species in some way, although I don’t make too much of an effort to look, and my species isn’t particularly mobile between patches. I would assume that this is because auto-evo cannot handle the preliminary work for the evolution of a nucleus. I don’t know if this is good or bad.
- Auto-evo’s energy and population prediction should be altered to account for the player’s cell size. Any additions to your cell will reduce your predicted population. With only the population number to look at, you’ll see that tiny cells are the only ones worth evolving, as being large reduces your population by an order of magnitude or more. This also means that you cannot tell from an auto-evo prediction whether adding a new part was actually good from your species, as there’s no way to tell apart the population loss from increased size from the population loss or gain from the part. I suggest dividing cell size by energy in the auto-evo prediction to account for this, but I don’t have enough knowledge to know if that’s actually a good solution, or if it would actually make you able to tell if the cell has improved despite its increased size.
- Toxins can be gravitated with vortex cilia. I’m not sure if this is intentional behavior or not, but it is rather weird. Gravitating toxin results in the toxins going everywhere when you turn off gravity. It’s the only way I know of to fire toxins in a direction that isn’t ‘forward’. Vacuoles also cannot store toxin, even though they can store mucilage. (They can’t store ingested matter as a specialty, but can as a generality, I believe, which is… bad.)
- Here’s a larger issue: MP BATTERIES. Modifiable parts like the cilia, pilus, slime jet, and toxisome have MP costs for their modifications. These MP costs store! If I change my toxisome to produce cyanide, that costs 8 MP. But if I, in another editing session, then switch it back to cytotoxin, I gain that MP back! This can put you above 100 MP. In the Multicellular stage, this technique gives you easy access to infinite MP. You can create a ‘battery’ cell type containing these parts (the slime jet is the most effective AFAIK, as its alteration costs 16 MP), and then duplicate it. In the new cell type, you can change the parts to gain their MP. Then, delete the new cell type. You have gained MP within an editor session. This means that, with patience, you can construct any multicellular organism in a single editing session.
- I know that the Early Multicellular stage is not a current priority, but through use of the above exploit, I found a good deal of issues with it. As mentioned earlier, multicellular organisms cannot rotate. That’s one glitch. The bigger issue is that if the size of your organism’s cells isn’t consistent (and probably even if it is), the hex structure completely disintegrates, producing organisms which utterly fail to match up to their design in the editor. There is a significant amount of empty space, and cells can get trapped inside your organism and be impossible to remove.
- Auto-evo in my run often cited the specific number of 394 steps, and got stuck on two specific numbers of steps. I don’t recall the exact numbers.
- For some positivity, phosphate chunks are a great idea. Ammonia can be produced by a cell, but phosphate can’t, so phosphate chunks even out the imbalance. (Actually, I find ammonia to be more commonly a limit now, with the prevalence of phosphate - this is because I don’t usually evolve to produce ammonia.) Also, the game handles large amounts of cells really well! My laptop isn’t quite a potato, but I didn’t buy it for its processing speed, and it doesn’t chug at all with so many cells on the screen. The increased abundance of cells in populated territories is really nice, although their distribution isn’t particularly even - you can swim through empty nothing and then see dozens of cells come into your screen. Also, auto-evo runs really quickly, and doesn’t require a long or even short loading screen anymore. Even in an advanced save, it didn’t require 1,100 steps or a full minute to load.
(I had to split these two because the forums wouldn’t accept the whole post, threw a 504 error.)
3 Likes
Congratz. You’ve reached the experienced Thrive player level as most players give feedback that being a predator is hard to play as, and doing iron is an “overpowered” strategy recommended to new players.
There was actually a bug where a patch would send too much of its excess compounds to a neighbour where that neighbour would end up with higher gas amounts (which is not really how an equilibrium would form). Fixing that was one of the fixes I included in my recent changes to the dynamic gases here:
What do you mean specifically?
From this description it sounds like the results are vastly different, which is excellent is it not? It would be super boring if auto-evo had consistent results between different playthroughs.
Are you making a statement? I’m 95% sure this is not the case. A tried mutation in the algorithm cannot be just a colour change, but any mutated variant can also have a colour change, so often when a species mutates it also gets a different colour.
Why? Each species in Thrive is given the chance to generate a bunch of mutations based on evolutionary pressures (as defined by miches) and the species will pick the best mutation it finds for itself. If there are many good mutations, one of them can be selected as a new offshoot species to split off, which then starts following the normal auto-evo logic from the next generation.
Which part of the GUI is this? I don’t recognize it from the terms you used.
Which tutorial is this?
The effect is rendered with world lighting meaning it is affected by the brightness of the current patch.
This is a guess but do you mean the chemoreceptor customization GUI? If not I’m again lost as to what you are referring to. But in the chemoreceptor GUI the compound amount slider is an advanced gameplay tactic that you don’t need to really touch (if it is not that I’m again totally lost as to what part of the GUI you are talking about).
The issue backlog strikes again:
I’ve known about the marine snow density not depending on existing species being a limitation since 2021, but again it’s one of those less visible things meaning that no one has volunteered to program a fix for this for over 3 years the issue has been open.
The prediction now primarily shows the energy gathered, which is unrelated to size. I really don’t want to remove the display for the predicted population as that’s effectively the life count in Thrive (how many times you can retry a life before losing the game). So while it would be possible to remove the population display entirely in the prediction if people complain too much, but I’m not really in favour of that.
It’s modelling a water vortex, so why wouldn’t toxins moving through the water be swept up by local currents?
This is a known bug:
I think you are describing this very often reported bug (which again no one is willing to program a fix for):
The new auto-evo miche algorithm is not split into as small steps as before. Especially the modify species step can take many seconds to generate the list of mutations to try for a single species, which is not optimal.
I think it was just the server having some issues still yesterday. Today it seems much better than the past few days.
Thrive’s variable enough that my thoughts on the easiest runs might be objectively incorrect since I don’t really go for ferrotroph runs. I think the driving reasons behind apex predation being easy for me are me playing with lower MP costs, vortex cilia being very good hunting weapons, and the gas glitches making mitochondria unbelievably powerful, so it’s not at all difficult to make a large, fast, quick-turning cell that can eat literally anything. At that point, the only threat is toxins.
Ingested matter specifically. Hunting while bound is already very difficult, and for some reason you essentially cannot accumulate reproduction in bound mode (is reproduction speed being divided by the number of bound cells?), but the ingested matter you get doesn’t work quite right when you unbind. I don’t fully, confidently recall, but either the ingested matter is shared among all cells, even those which don’t have anything engulfed, or the player cell gets the entire ingested matter quantity from the bound cell cluster.
I was speculating as to why auto-evo was producing identical divergent species, because the two single-cytoplasm cells I observed were identical in every way except color.
It seems more intuitively correct to me that, if auto-evo wanted to produce from Species A a Species B that had lost a part, and a Species C that retained it, Species B should be considered the divergence rather than Species A. A nomenclature issue, basically.
The GUI for variant parts, like the slime jet. When you open it, the box is much larger than the two options you’re given, and the size gives it the impression of being designed to have a maximum capacity of many more variants.
I don’t know the tutorial numbers or whatever you’re asking here, I just saw this tutorial pop up when I attempted leaving the editor without spending any MP.
It’s hard to learn advanced gameplay tactics if there’s no information about how they work. And I feel like one would think that the sensitivity slider would be important. I did, at least, when I was starting out.
Is it really unrelated to size? Didn’t know that. Thought it would be. Smaller cells get more energy because they’re lower on the food chain. Does a 1-thylakoid cell make the same amount of energy in the same patch as a 5-thylakoid cell would?
That’s totally intended, like how would matter physically teleport between cells? Each cell in a colony has their
individual ingested matter storage and ATP storage (as it would not be biologically accurate if ATP was shared).
No, but if you mean being in a non-multicellular colony. Reproduction progress pauses because it would be a total nightmare for the game systems to try to handle organelles dividing while in a colony. So that is not allowed. Also passive reproduction is on hold but I’m hoping this will be eventually fixed:
Can you share a screenshot at least? All ingested matter should still be tied to the cell that originally engulfed it. I foresee a potential bug where the player had nothing engulfed and after ejecting the colony the bar gets stuck displaying the previous value it had.
Being small is so beneficial that many species have convergent evolution. Which actually was highlighted as a really good indicator that the new system is better.
You do seem really into that idea. But making all the changes you’ve talked about like starting from the name generator etc. would take an extreme amount of development time considering that you are the first person to ever bring up this idea, whereas things have worked like this in Thrive for years. I’m not saying that it couldn’t be done but someone needs to volunteer a lot of programming effort for this to ever be really considered.
Ah the cell upgrade GUI. It’s by default a consistent size between different organelles. The tradeoff of needing to manually pick a size for each different organelle upgrade GUI and update that each time the number of options is changed, as well as less visual size uniformity in the editor, is a bigger problem than having extra empty space.
There’s a ton of new players who accidentally exit the editor without doing anything. So such a tutorial message is absolutely necessary to protect the player from themselves.
If I recall correctly the “staying small” tutorial recommends to stay small, and doesn’t say to not add anything.
The tutorial already says “most editor cycles” implying that it isn’t always required. I’m not sure how it could be made even more painfully clear without overbloating that tutorial message much more.
The official English text makes no mention of any “patch size”, it is like this:
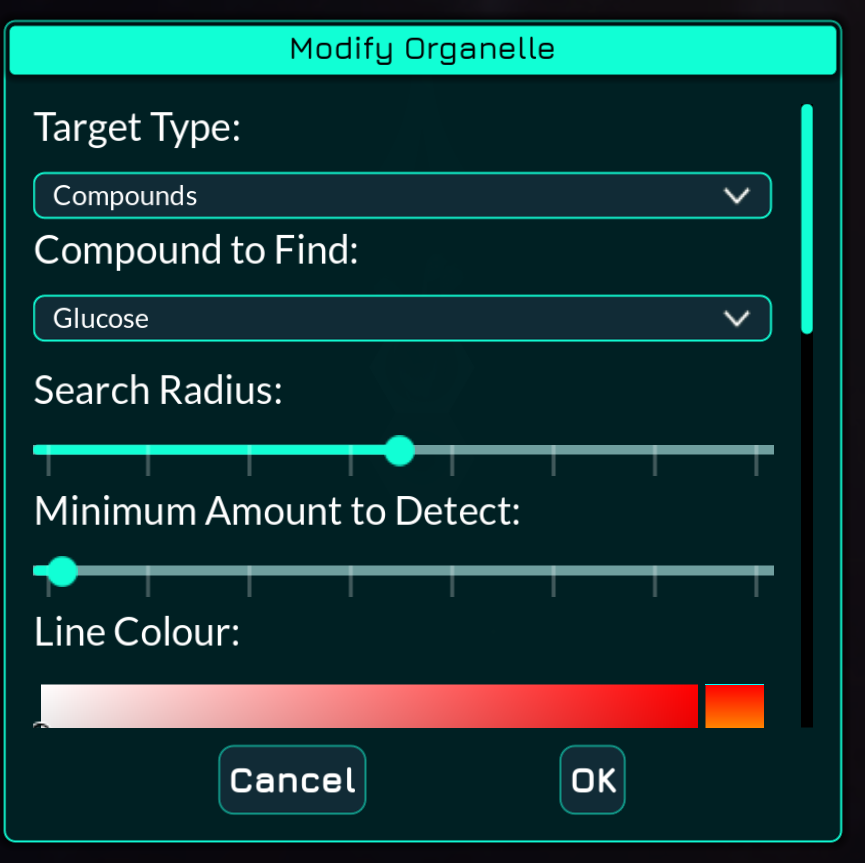
With my English reading skills it reads very naturally: “Compound to find is glucose, search radius is this, and minimum amount (of compound) to detect is this other slider”
I’m suspecting you are playing a translation of the game where the translator has not managed to preserve the clarity of the original text (and as such the translation should be improved).
Looking at those English text parts, even adding tooltips I would probably just write the same thing but as a full sentence like “Minimum compound amount to detect” and that again would be as clear if the translated text is the problem.
Depends entirely on how much energy it can gain from the miches it gets. For example I just tested and a simple cell with 1 thylakoid gets 177 energy, with two 304, and with 3: 304. So there’s optimal amounts of organelles to use to effectively compete for the limited energy that exists in a patch.
2 Likes
Still not very good at using my current Thrive computer, and also, it’s not the computer I write forum posts on. I’ll see if I can maybe get a screenshot at some point, whenever I play Thrive again.
That’s perfectly reasonable. What isn’t so reasonable to me is why cells don’t, say, evolve to have food parts above their cytoplasm instead of below. Convergent evolution for small body sizes still shouldn’t produce identical cells, statistically. A 1-food-part cell could show up in three different reasonable forms (the part above or below a cytoplasm, and just the part with no cytoplasm), but you usually see cyto above [food part] and, rarely, just the food part.
Also, all of these tiny cells that auto-evo loves don’t seem to show up in my lategame patches. Most of those patches are sparsely populated with large prokaryotes or offshoots of the player cell, at least in my recent experience.
Reasonable. I’m just concerned that a new player would get confused, because I heavily doubt this hypothetical new player’s intelligence. Which is basically like saying ‘teach this strawman how to play’, now that I think about it.
I play the English version. My complaint is that it’s not at all clear what the actual amounts on the ‘minimum amount to detect’ are. Are the ends of the scale ‘abundant’ and ‘very little’? Are they a narrower range? Does the search take into account the compounds in an entire cloud, or just a single tile’s worth of compounds?
I’m not attacking the ‘distance’ slider, even though it’s similarly unclear, because distance isn’t very relevant in Thrive unless you’re slow or out of ATP, and there’s no measurement in any case, so there would be no way to clarify that without adding a ruler.
Reasonable. It would probably be a good deal of work to make the UI scale to the buttons.
…
Now I’m wondering if Thrive’s already approaching the point where the system becomes too complicated and emergent to grasp. Would be an improvement, I suppose.
In an older playthrough I played a cell that was dependent on frantically swimming from glucose cloud to glucose cloud as quickly as possible. And I was highly dependent on the chemorecepter. The recepter seems to take into account the amount of surrounding compounds, the left being a slightest trace and the right being a whole lot.
I used maximum distance primarily in order to get the biggest range of view since having it sense the highest amout will make it only be able to detect a much smaller amount of clouds. Distance helps alot if you can only afford to move so far to find food.
I thought the sliders were perfectly intuitive, but perhaps adding images to each end would make it more clear.
The organelle placement algorithm tries to minimize the membrane size so it tries all positions radius 1 from the origin, then 2, then 3 etc. until it finds an empty position to place an organelle in. The order the directions are checked is consistent which is why the second organelle is placed in a consistent position. If someone wanted to they could add more randomness by for example randomizing the starting direction, and that would need just a few code changes.
This is much less likely to occur because the mutation algorithm would need to decide to remove the cytoplasm and place the other organelle in the same attempt, which is much less likely than just placing a new organelle.
Not even us developers know that without checking the source code. I can write some kind of more clarification tooltip but I won’t be able to add any kind of compound category markers. Though I can explain the second part that it detects individual compound points and not entire clouds.
This would be extremely hard to implement (like multiple times longer to do) so I can easily just add tooltips.
Good to know that I wasn’t completely incorrect in my assessment that the GUI is intuitive enough (and this has been basically the consensus for years as I think the upgrade is pretty old already).
3 Likes
…Chalk it into the ‘Easy Fixes, guys please…’ pile, then. For the second one - couldn’t you have the auto-evo algorithm consider cytoplasm an empty space? You can already place over it in the editor, and it’s not treated specially in any way - you don’t even get an MP cost reduction for putting a part on top of existing cytoplasm.
I can’t tell you how old the upgrade is, because 0.6.3 is the oldest version I’ve played, but it’s the only part I know of from that version which is modifiable. I didn’t test every part, but I wouldn’t be surprised if the chemoreceptor was the first part implemented to be modifiable - partly because, well, it wouldn’t be very good if it didn’t have that functionality.
2 Likes
I suppose that could be possible, but then that would mean that it isn’t really possible for auto-evo to place cytoplasm, which for example if this balance change goes through would be a major disadvantage: Glucose Consumption & Storage Balance Tweaks by Deus-Codes · Pull Request #5659 · Revolutionary-Games/Thrive · GitHub
So it’s again pretty complicated to make any change to auto-evo or code it depends on that actually makes things better.
I mean… this is specifically treating it differently. Back in the old days you couldn’t place anything on top of cytoplasm, but that was changed a really long time a go. And the way that works is that the placement logic prevents placing stuff when organelles overlap except if the overlapped organelle is cytoplasm. And the editor action logic has special storage space for replaced cytoplasms that it needs to put back if the action is undone. So there are a lot of special corner cases in the code for cytoplasm, and this has caused many bugs over the years.
Yeah, that’s probably the first organelle that was introduced that needed the upgrade system (unless I’m forgetting about some organelle).
3 Likes
Is this a good style of posting? I’ve got another massive batch of problems I noticed that I’ll post after I finish my no-engulf hunter run, and I’m wondering if I should put them here the same way as I did with these first two problem batches.
2 Likes
Well I think it is better than the alternative of posting one problem at a time and waiting for someone to reply before posting the next one.
2 Likes
So that would be a predator with a non-engulfation-allowing membrane, right? I can guess some things could be annoying about the way the chunks decay…
Turns out, it is very difficult to be a no-engulf hunter.
By which I mean it’s nearly impossible.
I didn’t really go too belgium the pili, but I should definitely have done so, because toxins are completely useless for hunting. They fire much too slowly. I kept on starving to death and had to drop the run.
2 Likes
Tbf all the walled membranes are pretty much meant for more sessile types of cells, not hunters.
1 Like
True. There are some cellular predators who don’t eat whole cells, I think, but they probably don’t have silicate membranes but they don’t have to eat cells. Basically all fungi are heterotrophs with chitinous membranes, not predators, but yes heterotrophic. They can usually turn into nigh indestructible spores and float until they get to a concentration of biomatter, which isn’t a feature in thrive, but yk.
1 Like
Well, obviously.
It’s still illustrative to me to see just how phenomenally terrible a hunter is if they can’t engulf. If you do this bad without engulfing as a hunter, then toxins and pili are basically useless as offensive adaptations. Slowdown toxins are still useful, but in a key-behind-the-lock fashion - they’re designed to counter fast cells, but fast cells will be too fast to be hit by the toxin. Pili require you to hit the enemy in melee, but this is difficult, since hunting necessarily has to be high-speed; you need to be fast enough to catch the enemy, and their main defense against predation is speed.
1 Like