This is a two-aparter of an idea, but they intermingle a lot in my head so I’m going to post them together.
Homeostasis
Homeostasis is a fairly self-explanatory concept; life likes things just how it likes them, and is generally very fickle to change (just think about how well humans do with too much/little oxygen). And, quite frankly, it’s unrealistic for a huge cell to just be able to freely dump waste and regulate all its internal components with no regard for its metabolism. Also, with thicker cell walls like a Calcium Carbonate Shell or a Doubled Membrane, it would be harder for the cell to dump waste.
That said, I think a cell’s metabolic processes should affect its internal chemistry. Gas levels, waste products, et cetera. This could also be used to “limit” a cell’s growth; as the cell gets bigger, it becomes less and less feasible to regulate its gas content; too many mitochondria will result in carbon dioxide poisoning, too many chloroplasts result in oxygen poisoning, and so on.
Example Scenario
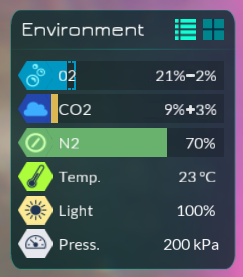
In the above image, the inverted colour segment in the Carbon Dioxide bar represents an excess of carbon dioxide, and the hollow segment in the Oxygen bar represents a lack of oxygen. Perhaps the cell is a metabolosome-dependent heterotroph of smallish-medium size; big enough to manipulate its internal chemistry, but not enough to truly screw up its homeostasis.
Chances are, you’d be able to view the overall chemistry in the editor; how much oxygen your cell produces/consumes, with the same going for nitrogen and carbon dioxide. At the very least, the net effect should be visible; rough estimates of your “working” chemistry.
That all said, of course, the concept of a hard limit is kind of… Well, bothersome, especially for those who are interested in making macroscopic, unicellular life.
That said, I believe that overall osmoregulation costs for organelles should be increased, however vacuoles should reduce osmoregulation cost (in animal cells, vacuoles and similar vesicles are used for waste disposal and transport all the time; if this would make vacuoles too overpowered, vacuoles could have to be designated for waste, making them cost ATP to run but providing the waste disposal properties).
Another concept, which would be a bit more intensive, is:
Osmoregulation cost could be accompanied by another variable called “regulation speed” which would be determined by your cell’s osmoregulation cost, individual units, and its resource absorption speed. Units that produce some obvious byproduct (Mitochondria produce Carbon Dioxide, Chloroplasts produce Oxygen, Rusticyanin produces Iron Oxide) would put “regulative stress” on your cell, which would have to be regulated by motile vacuoles or some similar regulatory unit.
The formula for regulation speed would probably be “osmoregulation cost^(1/2) * 100+(resource absorption speed)% + unit regulation speed boost” or something similar, where a cell’s size plays some role in determining its regulation speed (surface area) while providing diminishing returns due to the fact that volume scales faster than surface area (square-cube law, et cetera and et cetera).
Regulative Stress would be determined on a per unit basis. For example, mitochondria consume 1 oxygen and produce 1 carbon dioxide (numbers almost definitely not accurate), so their net regulative stress is 2. This, however, is where realism comes in to save the day:
Chloroplasts would consume 1.5 carbon dioxide and produce 1.5 oxygen, and so their effect compliments mitochondria’s: The net regulative stress of a system including bot 1 chloroplast and 1 mitochondria would be only 1, as the difference between their effects is 0.5 (positive or negative doesn’t matter, as having an excess and being starved both impede cell function).
Alternative Metabolic Routes
With all that talk of homeostasis, let’s have some fun with the potential for new resources and the metabolic pathways they could use. A small disclaimer: This is based less on earthly biology and leaning on the more sci-fi explanations of things. My personal issue with Thrive relying on earthly biology is that, although earthly biology is all we know, we can’t be sure there are cells that don’t, for example, electrolyze water to produce hydrogen and oxygen for respiration and chemosynthesis. So, take the biological implications of these next ideas with some grains of salt.
Hydrogen Sulfide Synthesis
This next subsection is dedicated to Hydrogen Sulfide synthesis, which includes 2 new resources and 2 new organelles:
-
Hydrogen would be a compound, however it wouldn’t be found in the environment; instead, the cell would be able to electrolyze water, expending ATP to increase both its Oxygen and Hydrogen levels.
-
Sulfur would be a mineral gradually released by iron chunks, allowing the player to harvest it like a normal compound.
-
The Electrolysis Plastid would be able to produce 2 hydrogen and 1% oxygen (for homeostatic purposes) via using 0.4 Iron and 8 ATP @ (room temperature) per second. (Seems low, right? ATP’s energy yield per mole is 30.8kJ, and water’s electrolysis requirment at room temperature is 237kJ per mole. Maybe I messed up the calculations, but it makes sense in my head.) Lore-wise, the Electrolysis Plastid uses iron at various ionic states to produce an electrictal charge, which is run through water to cause it to separate into gaseous hydrogen and oxygen. Electrolysis becomes easier at higher temperatures as water requires less energy to separate.
-
The Hydrogen Sulfide Synthesis Plastid is able to produce 0.6 Hydrogen Sulfide per second using 13 ATP and 0.6 Sulfur and 1.2 Hydrogen. Lore-wise, the Hydrogen Sulfide Synthesis Plastid (shortened to HSSP for convenience) is essentially a vesicle loaded with hydrogen sulfide synthesis proteins, catalyzing reactions that efficiently produce Hydrogen Sulfide with relatively low ATP cost. It keeps its contents heavily regulated, so exterior conditions are largely inconsequential.
The downsides for this system would be its energy and MP requirements to set up. That said, though, after all is said and done, Chemoheterotrophs would be able to survive in environments where Hydrogen Sulfide is naturally difficult/impossible to come by. The synthesis of Hydrogen Sulfide could also unlock symbiotic opportunities.
Radiotrophy
Another idea is the concept of Radiotrophy, where cells use melanin proteins to capture ionizing radiation and use it to drive a similar process to photosynthesis. This would include 1 new resource, 1 new environmental statistic, 1 new protein, 1 new vesicle, and 1 new organelle:
-
Uranium is a type of mineral, found in the form of small rocks that average 2-8 uranium when engulfed. Larger, unengulfable Uranium rocks, however, produce a green glow around them,
-
This is radiation, measured in Nanograys (1 Gray = 1 Joule per Kilogram of mass, so one nanogray means one picojoule per nanogram of mass, I think). Radiotrophs can laze around in this glow, using the radiation produced to power Radiosynthesis.
-
Melanin is a prokaryotic protein that uses radiation to produce glucose in a similar process to photosynthesis. Exact values can vary; perhaps 0.4 Glucose per second @ 9% Carbon Dioxide and 10 Nanograys (standard radiation levels from a medium-sized unengulfable rock at roughly 20 cell hexes away). One melanin provides protection against up to 3 Nanograys of radiation; if a cell is in too strongly irradiated of an area, it will take damage (1 unit of health per excess nanograys per second; for example, is the cell is in an area with 7 nanograys of radiation, but only has 3 radiation protection, it will take 4 damage per second).
-
The Melanin Vesicle is a eukaryotic vesicle (not an organelle, as it doesn’t have its own DNA) that acts as an upgraded version of the Melanin protein. It produces 0.9 Glucose per second @ 9% Carbon Dioxide and 10 Nanograys. One melanin vesicle provides a massive 10 Nanograys of radiation protection, making it the best source of radiation defense on offer. This is for more hyperactive players who want to be able to quickly recharge around rocks before going to zip around elsewhere.
-
The Radioplast is a eukaryotic, membrane-bound organelle that allows the cell to store uranium, up to 15 uranium per radioplast. Other storage increasing organelles do not affect the limit on uranium, as storing radioactive material unprotected is too dangerous. Radioplasts generate 0.6 Glucose per second using 0.01 Uranium @ 9% Carbon Dioxide per second. Being their own enclosed structures, they only provide a paltry 1 radiation protection. This is an option for those who want a reliable source of Radiosynthetic power they can take with them.
Starch: Advanced Sugar Storage
And the third idea I’m pushing today is more of a storage concept for autotrophs to take advantage of; during long periods where they are forced to rely on their stores, they are given a chance to store their sugars as even more compact forms. This idea contains 1 new compound, 1 new protein, and 1 new organelle:
-
Starch is a compound almost entirely exclusive to inside, the cell, though if forced to spawn in the physical map (i.e. if a cell containing Starch dies) it behaves like a mass of free-floating crystals, having to be engulfed to be consumed by the cell.
-
The Starch Weaving Protein is a prokaryotic protein that produces 0.3 Starch using 3 Glucose and 6 ATP per second if the cell has more than 75% of its glucose capacity filled with glucose. It can then reverse the process, producing 3 Glucose using 0.3 Starch and 6 ATP per second if the cell is below 30% of its glucose capacity. (This process also yields maltose in real life, which is used separately, however, I am unsure as to how accurate the devs want to go with this; I couldn’t find exact numbers for Glucose to Starch conversion, either).
-
The Amyloplast is a eukaryotic organelle that produces 1 Starch using 10 Glucose and 12 ATP per second if the cell has more than 75% of its glucose capacity filled with glucose. It can also reverse the process, producing 10 Glucose using 1 Starch and 10 ATP per second if the cell is below 30% of its glucose capacity. (Same disclaimer as the Starch Weaving Protein.)
And, well, that’s all I’ve got. Do I have more ideas? Maybe, but I spent 3 hours between research and writing on this post, so I’m calling this done for now.