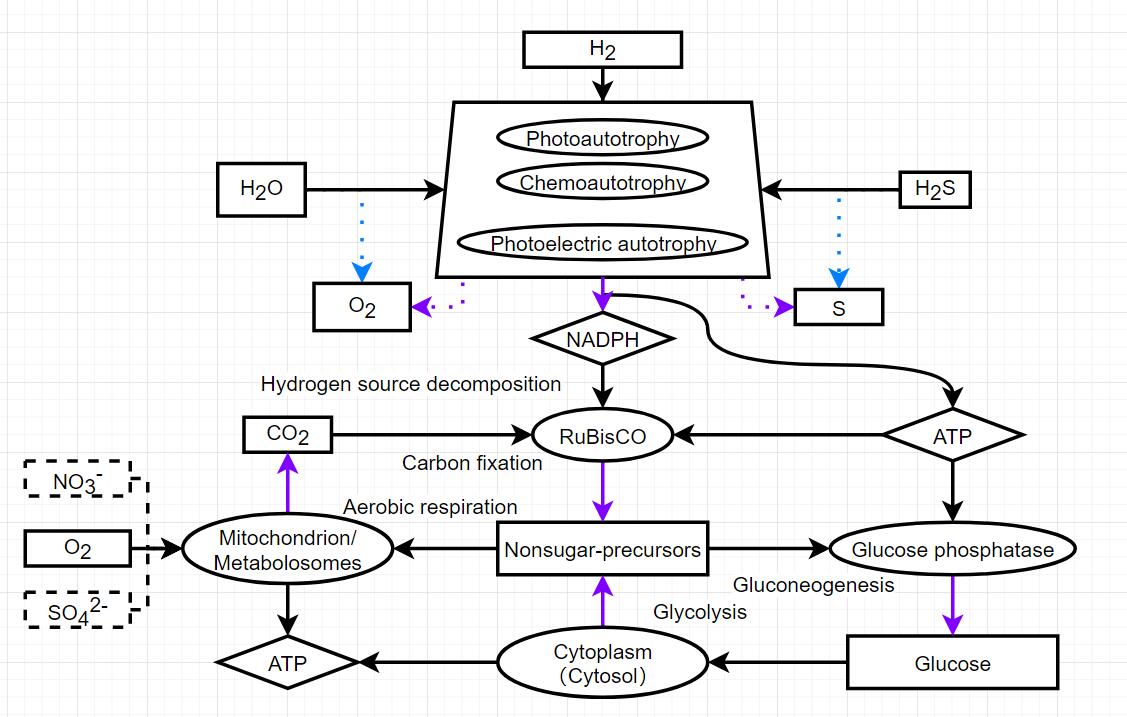
After a brief understanding of gluconeogenesis, I don’t agree to give up on RuBisCO. The problem of gluconeogenesis lies in the raw materials. It is not a carbon fixation pathway for CO2. Its role is to convert other non sugar organic substances. From the perspective of realism, it is necessary to store glycolytic metabolites (such as lactic acid, which may be better abstracted as nonsugar-precursors) as raw materials for gluconeogenesis. The conditions of action as a possible glycogen should also be good.
edit: Gluconeogenesis is the carbon utilization of non sugar organic matter by organisms, and we still need the carbon fixation way to obtain carbon dioxide.
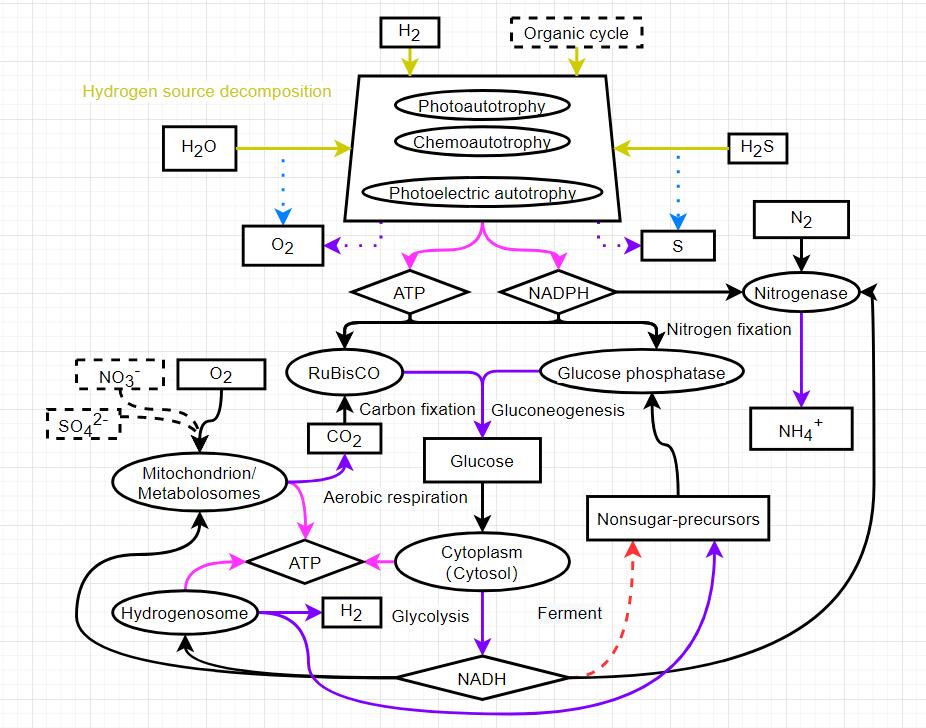
For the carbon sequestration process, I still prefer to use RuBisCO. As for its use in the game, I insist on splitting photosynthesis. Whether it is cyanobacteria or chloroplasts, thylakoid is just a place for light reaction, and carbon fixation is completed in the carboxysomes in cyanobacteria or chloroplast matrix, which is essentially the role of RuBisCO. At present, the direct synthesis of glucose by this thylakoid is easy to misunderstand that thylakoid can independently complete photosynthesis. Perhaps the evolution of RuBisCO predates photoreaction and was born when cells first used hydrogen as a hydrogen source for carbon sequestration. Not only photosynthesis, but also various chemoautotrophic processes rely on RuBisCO for carbon sequestration, and this commonality should be reflected.
Note: NADH indicates the degree of glycolysis and can be regarded as NADH and the corresponding amount of pyruvate. NADH allows incomplete utilization and all excess NADH disappears. It can be considered that the disappeared NADH and pyruvate enter the fermentation pathway.
In my imagination, NADH and pyruvate produced by glycolysis will enter anaerobic respiration by default if they are not further utilized, which does not require the placement of special organelle.
As for the types of anaerobic respiratory products, I am not yet aware of the effects of different types of anaerobic respiratory products on cells. May the type of product be determined by the passive protein system? Different anaerobic respiratory product enzymes have different shapes when placed (puzzle placement of passive proteins).
Excess Nonsugar recursors will be excreted like excess substances from digestion now.
Unfortunately, developers seem to be very resistant to adding more compounds, as they are concerned that this will make the game more complex. In my opinion, for a realistic simulation game, one should not avoid the complexity brought about by reality, which is itself a part of gameplay. For players who pursue realism, these complexities are nothing at all. One advantage is that this game will gradually become more complex from a simple environment, and players’ cells can also explore complexity from simplicity, while maintaining simplicity is not an inability to survive. This will bring diversity.
So I hope the splitting process can bring modularity and balance control, so that players can get small enough parts. This will bring exploratory depth to the game.
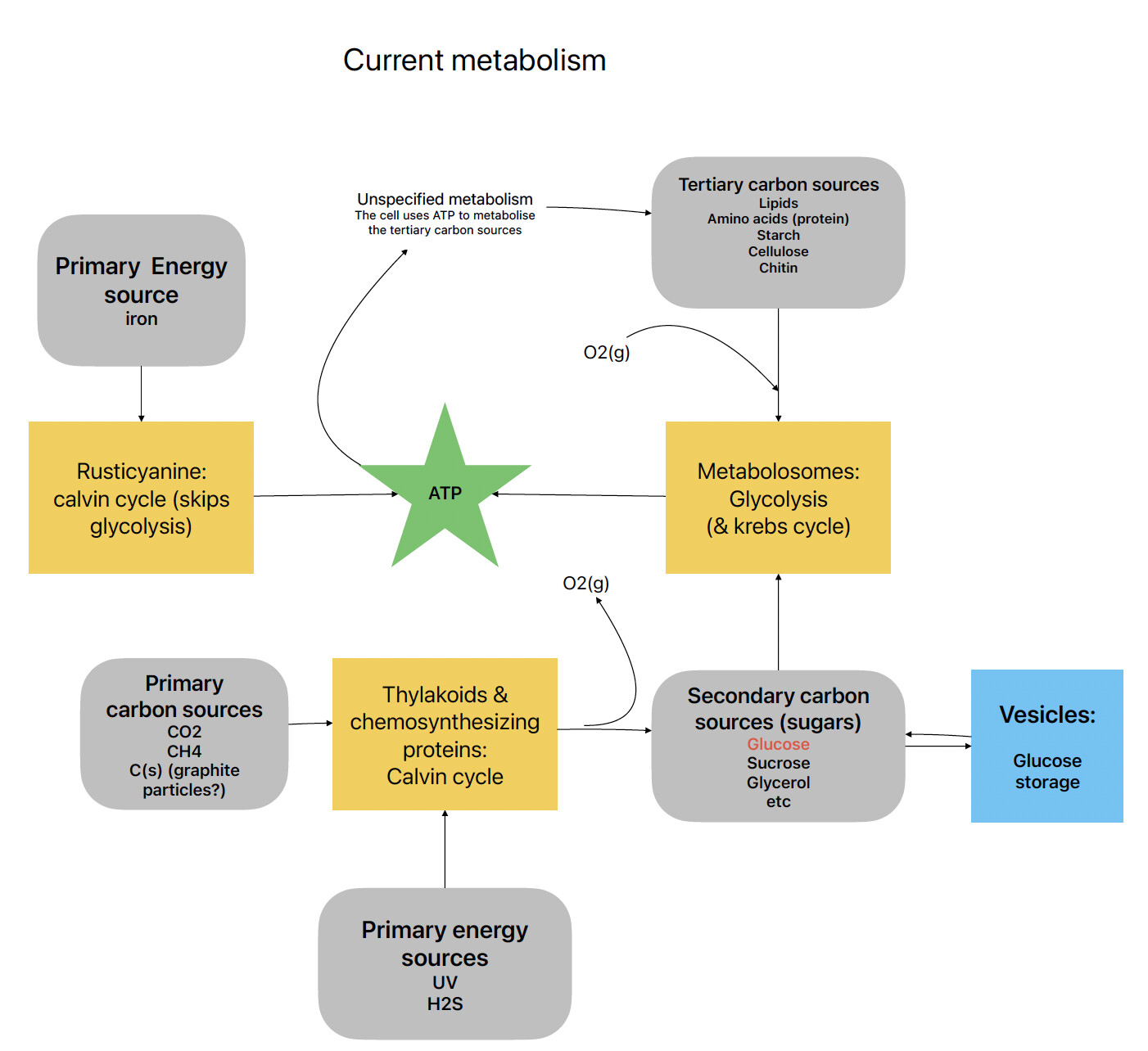
I completely agree to not give up on remodeling the metabolism and gluconeogenesis, especially because rustacyanine kinda sucks right now. Gaining glucose from other sources of carbon would better illustrate how to think of metabolism in real life.
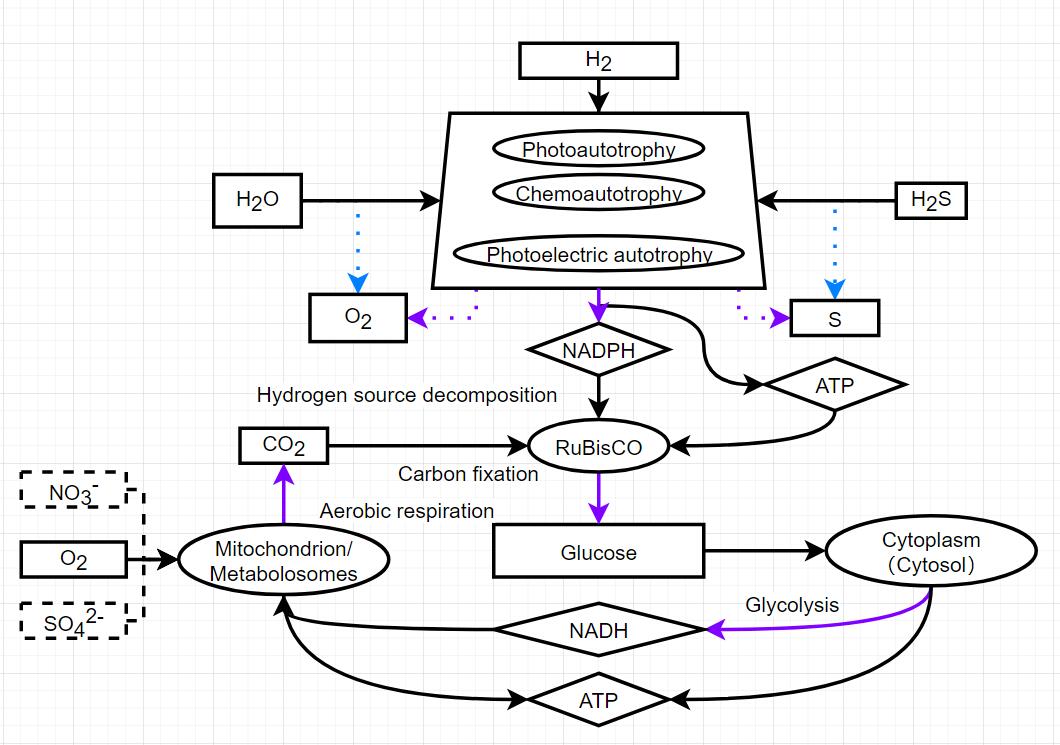
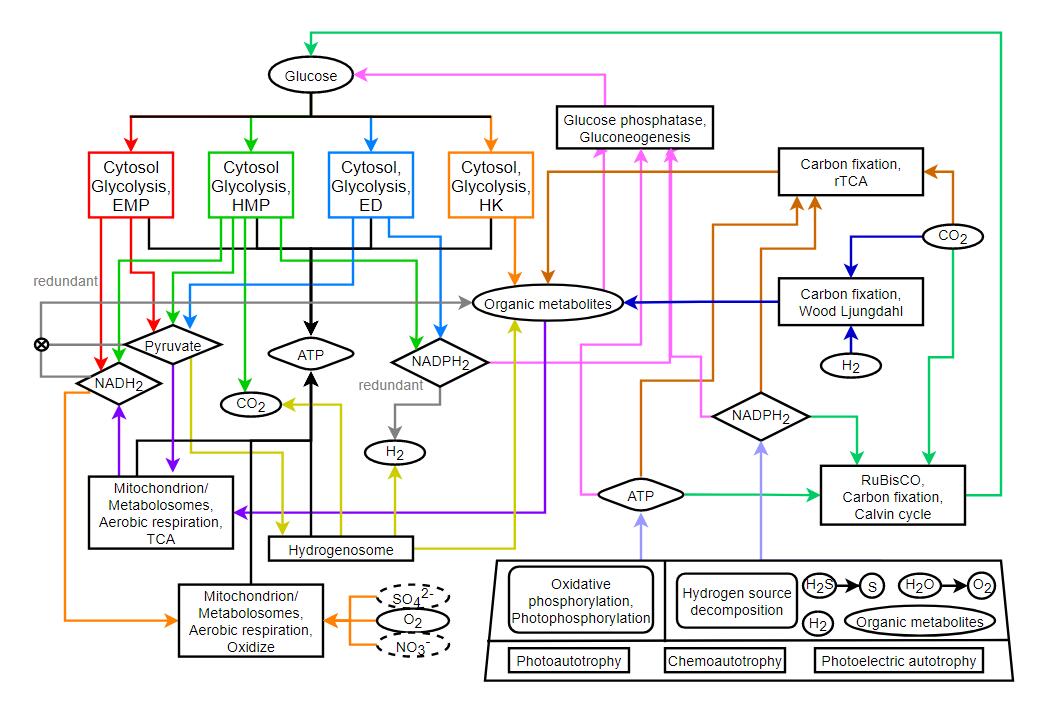
I made a mock up of a much simpler version of the graph you made above that is both more accurate than the current game while being less complicated (detailed), and I think it will also help balance the different survival strategies very well. Trying to make a more realistic pathway is always a possibility, but can very quickly become too complicated, especially when you want to move on to later stages in the game (multicellular). All of the organelles and metabolic strategies would have to be taken into account, this includes all enzymatic pathways.
.
Separating carbon fixation and other sources of energy (light, radiation, iron and hydrogensulfide) just makes sense and will actually help balance the game better. After getting a metabolosome, you could gain rustycyanine first, then a RuBiSco to store any energy gained from iron into glucose. This allows your little cell to venture outside of the iron cloud without their ATP draining instantly which is what currently happens. This has been implemented as a native part of thylakoids and the chemosynthesizing proteins, but separating the two processes would make sense. To prevent players from gaining thylakoids or rustycyanine without a RuBisCo, you could grey out the thylakoids until a rubisco is part of the cell.
i feel like disabling harvesting energy from light until you can turn that energy from one biologically available form to another wouldn’t exactly make sense but that’s a good diagram
I mean, in every biome you start out with glucose clouds so you don’t need a thylakoid until at least your second division. The thylakoid could just generate ATP like the rustycyanine does currently. Then by including RuBisCo, you gain a way of storing the energy harvested from the sun in the form of glucose. This would be a lot more accurate to how it works IRL, and I think it would be much easier to implement into the game. The diagram was for improving the game with the logic it has now. Making it any more complicated would require a massive overhaul which is why I suspect the Devs are resistant to making any of these changes.
As for glluconeogenesis, this would only be necessary further down the line when you want your cell to specialize in heterotrophy (they eat organic sources of carbon, or according to my diagram tertiary carbon sources).
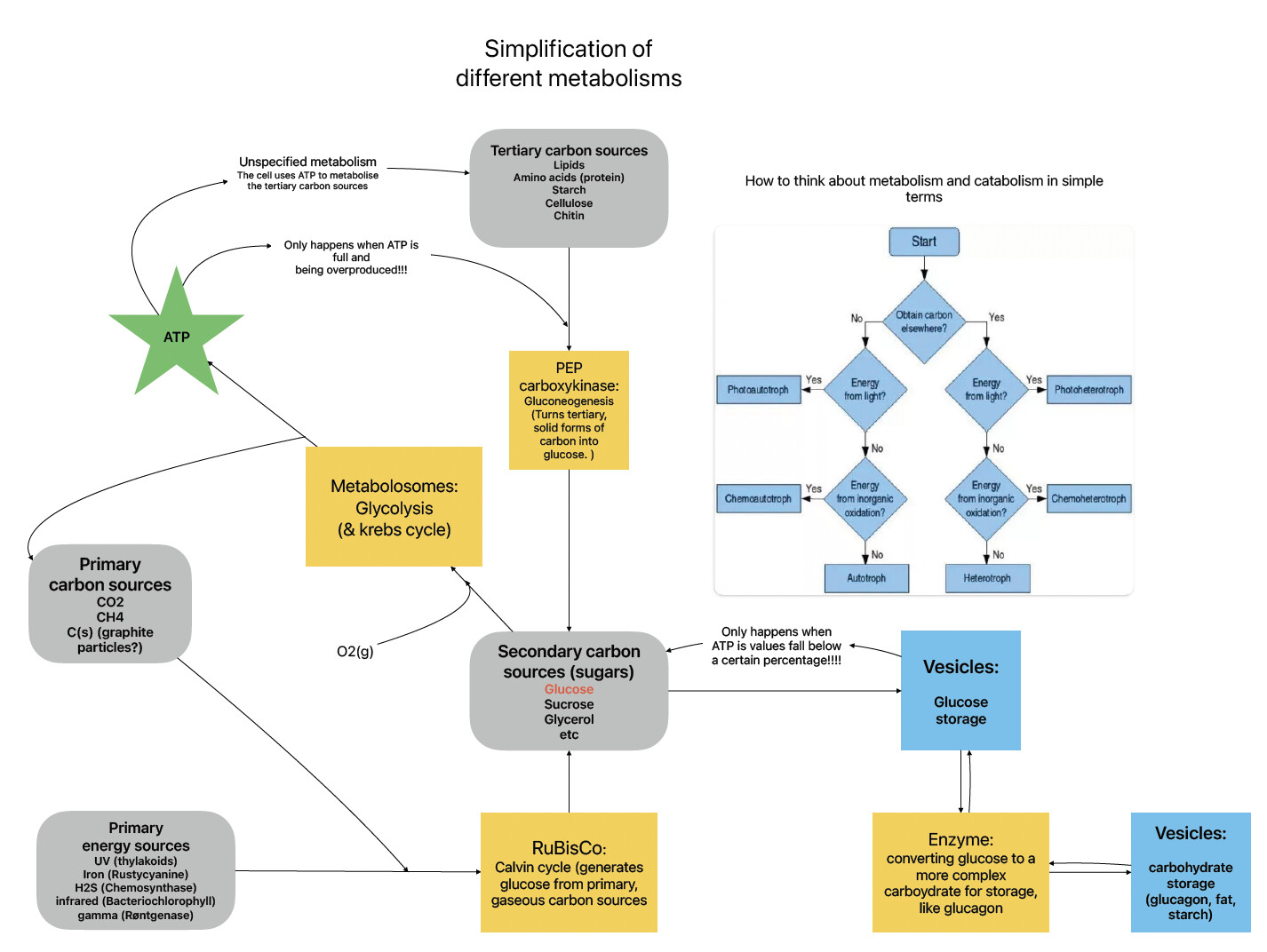
The biggest problem is to avoid consuming glucose to synthesize glucose. That’s why I want to add [NADPH], which will become a gauge of autotrophic intensity to limit the input of ATP for glucose synthesis.
The initial version of each autotrophic organelle is pure ATP production, with NADPH serving as an upgrade to the organelle.
As for glluconeogenesis, if pyruvate is added, my idea is to allow cells without carbon fixation ability (retinol phototrophic archaea) to synthesize pyruvate into glucose when glycolysis does not occur.
The third level carbon source is not necessary, they are in a sense included in digestion. If a newly created substance is only used to store glucose, it is better to directly add it to specialized stored organelles.
For example, fat droplets and starch granules, specialized glucose storage.
We essentially agree on everything. Especially the autotrophic organelles working as a primary energy source(whether that’s ATP or something else to simplify the implementation). As for carbohydrate storage: you could have specialized “vesicles” that can store the different carbohydrates for now just by modifying the vesicle when you add it to your organism. Kinda like how lysozyme works rn.
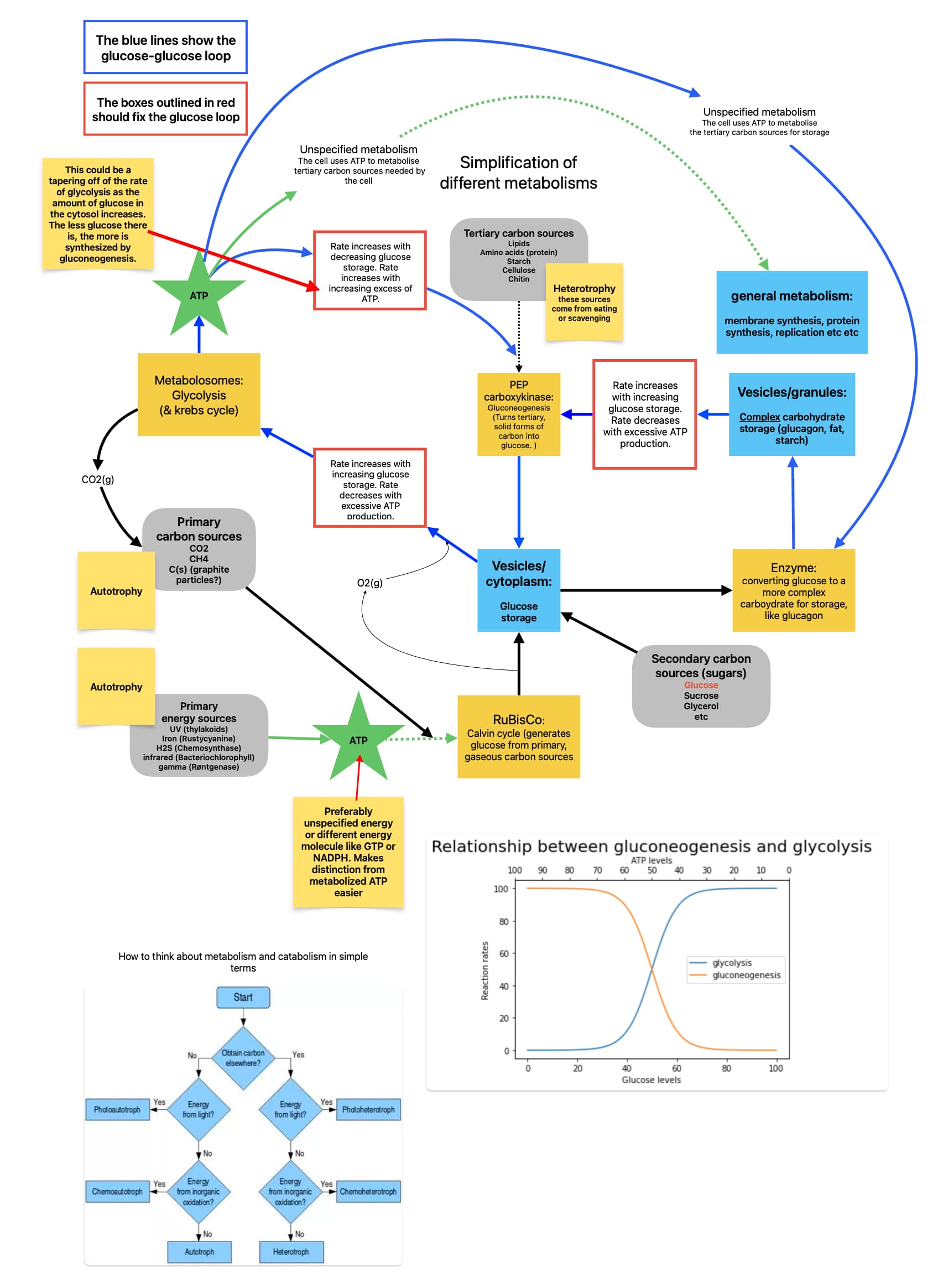
I think that there is a way to stop the cycle without pyruvate, and the only reason I’m trying to make a decent argument for this is because it would be a much easier step in the right direction. If it works then the later addition of pyruvate and NADH might make the whole system even better and more stable. But showing these diagrams to non-biologists/biochemists is kinda overwhelming, and I think just difficult to code to start out with (I’ve had to code systems like this before for courses and they quickly turn into a nightmare). Gotta build a solid foundation both theory and programming-wise, then move up from there.
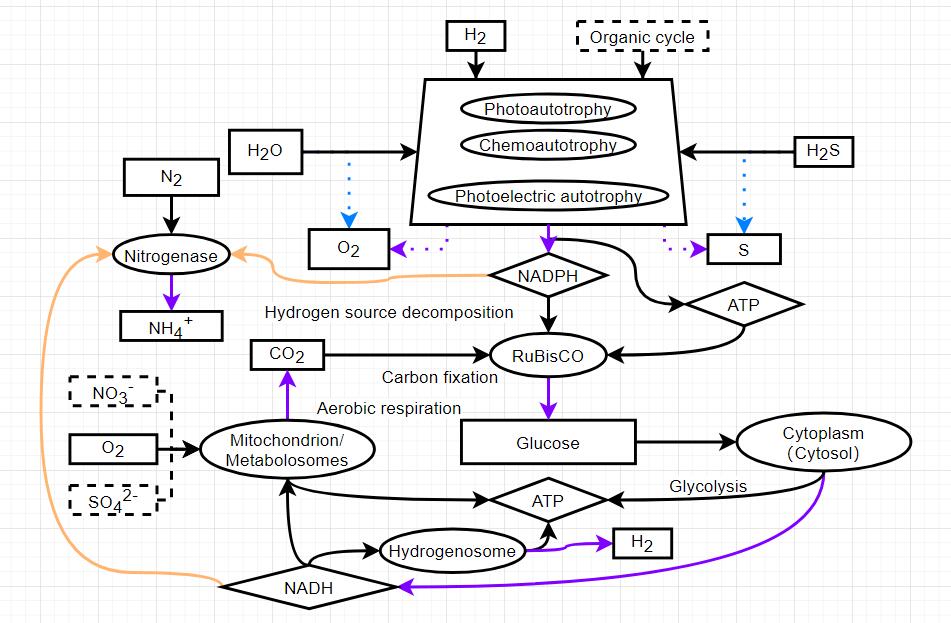
Here’s a new diagram showing an auto-inhibitory pathway for gluconeogenesis and glycolysis. I could be missing something though. Please point out any hole in this mechanism so that I can improve it! If I haven’t stopped the loop please explain how
I mean, you ARE interested in a pretty nerdy game. I hope the game helps you achieve that goal! It’s definitely making my own studying for my thesis much more fun! The more diagrams you see of metabolic pathways the better equipped you’ll be for biochemistry.
Do you seem to want to use glucose storage as a scale?
The biggest problem is that the current ATP consumption in games is only different between stationary and mobile, and the metabolic process is relatively fixed. At present, the storage of games is extremely overflowing, making it difficult to assess the impact of glucose storage.
I’ll take a look at the link when I have time but I think you’re right. It looks interesting and I’ve wondered the same things.
Glucose storage as a scale for both glucose and energy production and consumption, yes. Just like irl where there are many auto-inhibitory metabolic reactions, where the product of a reaction inhibits its own production. This prevents an overproduction of the product and limits the energy resources used in creating it. In fact, the main "tarting enzyme for glycolysis hexokinase is autoinhibited by its own product glucose-6-phosphate, ensuring that ATP doesn’t go to waste and that blood sugar levels don’t drop. This mechanism has other important pathways that it regulates (g6p is an important substrate in other pathways), but for an oversimplified game-model, I think it can regulate glycolysis.
"Hexokinase and glucose transport
Hexokinase is nominally the first step in glycolysis, but it is not the major point of regulation because glucose 6-phosphate is a branch point leading to glycogen and to the pentose phosphate pathway. Importantly, glucose 6-phosphate is an inhibitor of hexokinase, so if the other pathways are slow and if phosphofructokinase is inhibited, then glucose 6-phosphate will increase and inhibit hexokinase." source
So, if I understood you correctly, then I think glucose storage could be scalable depending on your cell’s size and growth rate (and by extension metabolic rate). This would hopefully fix that part of the realism.
And I really do think more realism would be really cool, but if you look at one of my favourite images of all time you’ll see why this can very quickly spiral out of control. Often an easy (and less realistic) fix will be better for a game.
So I think controlling biological processes through storage is too difficult, and I prefer to control the source and consumption of ATP.
Even in reality, controlling the speed of biochemical reactions and whether they occur is based on the state of the enzyme and the level of substrates and products. But for games, we only have environmental conditions and ATP consumption. Games require steady state rather than constantly adjusting dynamic balance in reality.
then you know more about game programming than me. I have simulated similar systems (in python) for school projects in chemistry with the concentrations of different ions in different water reservoirs depending on the rainfall and evaporation rates as well as the overflow from one reservoir to the other and out into the rivers. Kinda the same concept but not as complicated and not part of a game.
Another option for now, is to merely have all of the structures that capture energy for the calvin cycle to generate glucose directly and for metabolosomes to “digest” organic matter, converting it to glucose and the other elements available in the game rn. this is exactly what happens with most structures in the game rn anyways. I don’t think its the most elegant solution, but it does avoid changing the rates of different reactions as well as a massive overhaul of the mechanisms in the game.
In fact, a simpler way is to use an upgrade system that allows organelles to switch between focusing on ATP production and glucose production, which should be achievable now.
But I still want to see them become separate processes, carbon sequestration processes that exist relatively independently in organisms. Carboxysome is an organelle like structure that can concentrate CO2 from the environment
Imagine that there are three forms of autotrophic organelles:
Focusing on ATP production (primitive autotrophic);
Production of ATP and [NADPH] (calvin cycle, advantage at low concentrations of CO2);
Production of glucose (rTCA, advantage at high concentrations of CO2).
Transforming organelles between the ‘1,2’ forms does not consume MP, and the MP cost of placing Carboxysome (carbon fixing organelles) is relatively low compared to the ‘3’ form, such as one Carboxysome satisfying three thylakoids.
rTCA has the advantages of small size and high concentration of CO2, making it more suitable for the chemical autotrophy of the seabed;
Calvin cycle has the advantages of saving MP and low concentration CO2, making it more suitable for surface photosynthesis. You can also see it in large cells on the seabed.
Hahaha, I’m trying to make a “simple” model in python and I’m starting to realize why pyruvate NADPH and NADH might be necessary lol. Does the dark side still have cookies? XD